
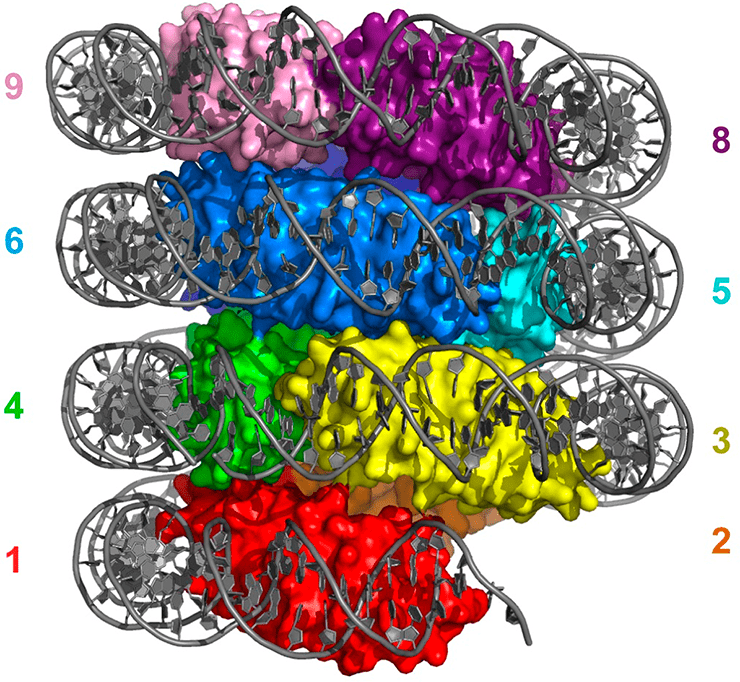
Модель гипернуклеосомы архей на примере гистона HMfB. Димеры гистона формируют левозакрученную спираль, на которую намотана ДНК. Разными цветами отмечены разные димеры гистонов, ДНК закрашена серым.
Археи, несмотря на то, что не имеют оформленного ядра, по очень многим признакам гораздо больше похожи на эукариот, чем на бактерий. В частности, их геномная ДНК упакована и компактизирована с помощью гистонов, как у эукариот. Однако гистоны эти весьма своеобразны (как, наверное, и всё у архей): в отличие от гистонов эукариот, они не формируют стабильные октамерные нуклеосомы, хотя третичные структуры гистонов архей и эукариот очень похожи. Последние исследования свидетельствуют, что «нуклеосомы» архей не имеют фиксированного размера и состоят из различного числа димеров гистонов, причем плотность упаковки ДНК с помощью таких вариабельных нуклеосом напрямую связана с репрессией транскрипции связанного с ними участка ДНК. Что наиболее удивительно, длина нуклеосом архей, похоже, может быть практически неограниченной, за что исследователи назвали их гипернуклеосомами. Впрочем, с помощью биоинформатического анализа у некоторых архей удалось найти гистоны с сильно отличающейся от остальных аминокислотной последовательностью, которые, по-видимому, неспособны формировать гипернуклеосомы. Наконец, у некоторых архей есть гистоны с N- и C-концевыми хвостами, которые похожи на хвосты гистонов эукариот и тоже могут подвергаться посттрансляционным модификациям. Так каковы же они, гистоны архей, и как устроен хроматин архей? В статье мы постараемся ответить на эти вопросы.
Необходимость поддерживать пространственную архитектуру хроматина стоит перед представителями всех трех доменов жизни: архей, бактерий и эукариот. У бактерий имеются особые белки — нуклеоид-ассоциированные белки (nucleoid-associated proteins, NAPs), позволяющие изгибать или переплетать спирали ДНК, и эти белки не только определяют структуру бактериального хроматина, но и влияют на активность определенных участков генома. К числу таких белков относятся, в частности, гистоноподобный белок HU из штамма U93 E. coli, а также H-NS — гистоноподобный белок, структурирующий нуклеоид. У эукариот и большинства архей задачу упаковки геномной ДНК выполняют гистоны. Хотя наша статья посвящена гистонам архей, будет нелишним напомнить, как устроены гистоны эукариот.
Гистоны эукариот
У эукариот ДНК, подобно нити, наматывается на «катушки» — нуклеосомы, состоящие из восьми молекул гистонов: по две молекулы гистонов H2A, H2B, H3 и H4 (рис. 1). Строго говоря, нуклеосома состоит из двух димеров H2A—H2B и тетрамера H3—H4, на которые намотана ДНК длиной около 146 пар оснований двумя витками. Для всех гистонов характерен особый структурный мотив, известный как гистоновая складка (histone fold): две короткие и одна длинная α-спираль, разделенные петлями. Существуют предположения, что в некоторых условиях, например, при элонгации транскрипции, функциональную роль начинают играть «неканонические» комплексы гистонов: тетрасомы, представляющие собой тетрамеры H3—H4, гексасомы (тетрамер H3—H4 и один димер H2A/H2B), а также гемисомы — димер H3—H4 плюс димер H2A/H2B. Помимо кóровых гистонов, у эукариот имеется также линкерный гистон H1, лишенный гистоновой складки. Он взаимодействует с ДНК прямо в точке ее входа на нуклеосому и выхода с нее. Важную роль в компактизации ДНК эукариот играют N-концевые хвосты гистонов, которые могут подвергаться многочисленным посттрансляционным модификациям — ацетилированию, метилированию, фосфорилированию, убиквитинированию и биотинилированию, что имеет колоссальное значение для регуляции экспрессии генов.

Рисунок 1. Сравнение гистона и гистонового октамера эукариот и гистона архей. а — Эукариотическая нуклеосома. Желтым цветом окрашены гистоны H2A; красным — H2B; голубым — H3; зеленым — H4. б — Архейный гистон HMfB. HMfB — histone B из Methanothermus fervidus.
Ноу-хау архей: бесконечная гипернуклеосома
Что же касается архей, то гистоны имеются у большинства видов в самых разных группах. Любопытно, что у некоторых есть и гомолог бактериального гистоноподобного белка HU, а у многих архей идентифицированы особые высококонсервативные нуклеоид-ассоциированные белки семейства Alba, которые, впрочем, найдены и у эукариот (рис. 2). Эти белки способны к формированию филаментов при взаимодействии с ДНК, а также соединяют дуплексы ДНК подобно мостикам. Эксперименты в условиях in vitro показали, что связывание ДНК с белками Alba подавляет транскрипцию. Любопытно, что у большинства архей типа кренархеотов отсутствуют гистоны, и ведущую роль в организации хроматина играют NAPs.

Рисунок 2. Модель организации хроматина архей
Существуют и другие, более малочисленные белки, влияющие на упаковку ДНК, которые содержат некоторые кренархеоты и эвриархеоты. Например, у кренархеотов имеются специфические мономерные белки, изгибающие ДНК, — Cren7 и Sul7.
Гистоны архей, как и гистоны эукариот, имеют гистоновую складку, однако, как правило, лишены N-концевых хвостов. Не выявлено у архей и линкерных гистонов, гомологичных эукариотическому гистону H1.
В растворе гистоны архей образуют димеры — как гомодимеры, так и гетеродимеры, поскольку многие археи синтезируют гистоны нескольких видов. К примеру, архея Methanothermus fervidus производит два разных гистона, уровень синтеза и относительное соотношение которых различны в разных фазах роста, поэтому, возможно, эти белки имеют разное функциональное значение.
Имеются сведения, что как in vitro, так и in vivo гистоны архей могут связывать ДНК в виде тетрамеров. Ряд исследований показал, что в действительности ДНК архей связана с неограниченным числом димеров гистонов, которые, взаимодействуя друг с другом, формируют ДНК-белковый филамент вариабельной, теоретически неограниченной длины. Кристаллографическое изучение гистона HMfB вышеупомянутой археи M. fervidus показало, что in vitro его молекулы формируют бесконечную левозакрученную нить — гипернуклеосому, — причем чем больше витков в ней, тем плотнее упакована намотанная на нее ДНК. Когда структуру гипернуклеосомы у археи Thermococcus kodakarensis экспериментально нарушали, вводя мутации в гистоны, архея начинала иначе реагировать на определенные питательные вещества, что свидетельствует о роли упаковки ДНК архей в экспрессии генов.
Изучая гистон HMfB, исследователи выяснили, что он образует димеры за счет гидрофобных взаимодействий аминокислотных остатков, расположенных в глубине молекулы, а также посредством гистоновых складок, для формирования которых необходим один крайне важный солевой мостик. Эти гидрофобные остатки, а также остатки, образующие солевой мостик, оказались высококонсервативными среди гистонов архей, поэтому гистоны архей, вероятнее всего, в большинстве своем имеют схожую структуру. Кроме того, консервативными оказались и аминокислоты, которые обеспечивают взаимодействие гистонов архей с малой бороздкой ДНК. На основании перечисленных данных высказали предположение, что гипернуклеосомы характерны для хроматина большинства архей. Впрочем, некоторые новооткрытые археи из прежде неизвестных типов имеют крайне необычные гистоны, о которых расскажем ниже.
Эволюция гистонов
Предполагается, что эволюционно гистоны эукариот произошли от гистонов архей. Родство этих белков подтверждается высоким сходством и аминокислотных последовательностей, и вторичных структур. По расчетам исследователей, первые гистоны появились более одного миллиарда лет назад именно как структурные белки, а участвовать в регуляции экспрессии генов они стали гораздо позже, когда у эукариот обособились четыре типа коровых гистонов. Интересно, что гистоновая складка есть не только у некоторых эукариотических белков, не являющихся гистонами (таких как некоторые факторы транскрипции), но и у белков некоторых бактерий. Биоинформатический анализ последовательностей этих белков показал, что мотив гистоновой складки существовал уже во времена последнего универсального общего предка (LUCA).
Как упоминалось выше, элементарная единица комплексов гистонов большинства архей — димер, но у архей класса Halobacteria имеются необычные тандемные гистоны. Эти гистоны представляют собой сшитый в одну полипептидную цепь димер с двумя взаимодействующими гистоновыми складками, что подчеркивает важность и необходимость гистоновой складки для формирования такого димера. Вместе с тем давление отбора на остальную часть молекулы тандемного гистона ослабевало, и, согласно одной из гипотез, из тандемного гистона в конце концов появился асимметричный гистон-димер, который мог стать предком эукариотического димера H3—H4. Впоследствии асимметричный гистон разъединился на два разных белка, кодируемых двумя разными генами. Действительно, тетрамер гистонов архей структурно очень близок к эукариотическому тетрамеру H3—H4, а гистоны H2A и H2B, как предполагается, произошли позже от разделившихся гистонов H3 и H4. В пользу такого сценария свидетельствует и тот факт, что гистоны H3 и H4 гораздо больше похожи на гистоны архей, чем гистоны H2A и H2B.
Далее у эукариот начали появляться так называемые неканонические формы гистонов, которые часто приурочены к определенным клеточным процессам, причем, что интересно, неканонических форм гистонов H2B и H4 практически неизвестно, в то время как для H3 и H2A они описаны в изобилии. Дальнейшим усложнением эукариотических гистонов стало формирование октамеров, на которые ДНК наматывается в виде двух витков, а не одного, а также появление N-концевых хвостов, которые позволили еще плотнее компактизировать ДНК и стали подвергаться интенсивным посттрансляционным модификациям.
Но наиболее интересно обстоит дело с гистонами у новооткрытых архей, которые стали известны науке благодаря бурному развитию метагеномики . Например, в собранном геноме LC_3 предполагаемого типа Heimdallarchaeota удалось найти гены целых десяти гистонов — абсолютный рекорд для архей! Вместе с тем у представителей предполагаемых типов Parvarchaeota, Geothermarchaeota и Vestraetearchaeota гистонов не нашлось вовсе: у этих архей, по-видимому, за компактизацию хроматина отвечают NAPs, тем более что у некоторых из них удалось найти упаковывающие белки вышеупомянутого семейства Alba. Похожая ситуация и в случае относительно давно описанных кренархеот, подавляющее большинство которых не имеет гистонов. В геномах Geothermarchaeota и Vestraetearchaeota исследователи пока не смогли найти генов, белковые продукты которых походили бы на известные белки хроматина. Наконец, у представителей предполагаемого типа Thorarchaeota есть ген, кодирующий белок HU (как мы помним, он отвечает за упаковку ДНК в нуклеоидах кишечной палочки).
Посттрансляционные модификации гистонов архей
Долгое время было принято считать, что гистоны архей не подвергаются посттрансляционным модификациям, а их регуляторная роль сводится к блокировке транскрипции ДНК, с которой они связаны, за счет того, что они не дают РНК-полимеразе II связываться с ДНК и синтезировать РНК-транскрипт. Так, у археи T. kodakarensis, имеющей два разных гистона, делеция одного из них приводила к почти десятикратному усилению экспрессии около 5% генов (однако делеция обоих гистонов оказалась несовместима с жизнью, что указывает на важнейшую роль гистонов в функционировании клетки). Ранние эксперименты по транскрипции in vitro с транскрипционной машинерией археи Methanothermobacter thermoautotrophicus показали, что добавление гистонов к раствору с ДНК и белками транскрипции снижало количество вырабатываемых транскриптов, а при добавлении гистонов в концентрации, в которой они, согласно расчетам, должны связывать всю находящуюся в растворе ДНК (около 1 димера гистонов на 30 пар оснований ДНК) синтез РНК останавливался вовсе. Схожие данные были получены для транскрипционного аппарата Pyrococcus furiosus.
Считается, что архейные транскрипционные факторы TBP и TFB способствуют транскрипции, поскольку конкурируют с гистоновыми димерами за связывание с ДНК и в конечном счете вытесняют их (рис. 3а). У T. kodakarensis удалось выявить, что фактор транскрипции TFS, а также комплекс Spt4/5 облегчает взаимодействие РНК-полимеразы с ДНК, что также активирует экспрессию генов (рис. 3б).
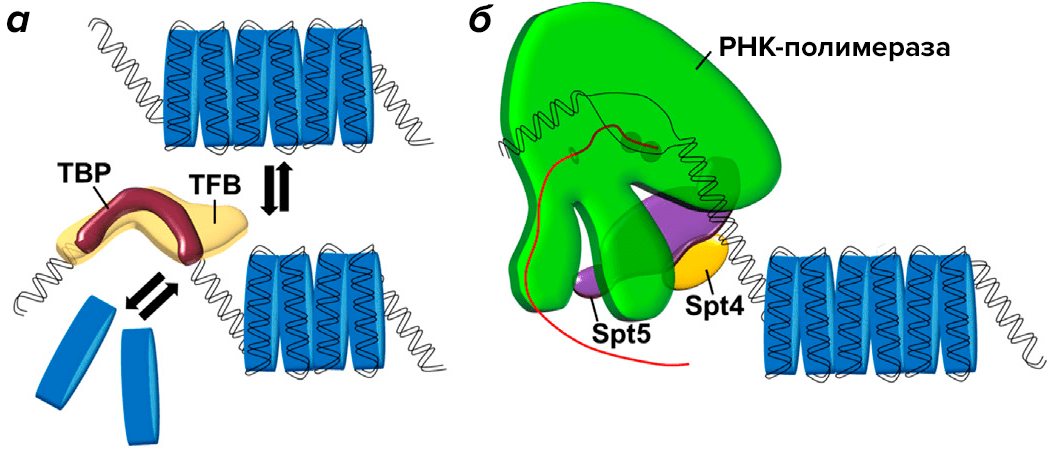
Рисунок 3. У архей транскрипционные факторы вытесняют димеры гистонов с ДНК и тем самым способствуют активации экспрессии генов
Новооткрытые группы архей принесли биологам и другие сюрпризы. Оказалось, что некоторые представители предполагаемых типов Heimdallarchaeota, Huberarchaea и Bathyarchaeota имеют гистоны с N-концевыми хвостами, как гистоны эукариот! Хвосты двух гистонов, найденных у Heimdallarchaeota и Huberarchaea, имеют примерно ту же длину и аминокислотный состав, что и хвост гистона H4. Поскольку четыре остатка лизина в N-концевом хвосте H4 подвергаются ацетилированию, гипотетически они могут ацетилироваться и в хвостах гистонов архей, тем более что в их геномах найдено несколько предполагаемых лизинацетилтрансфераз и деацетилаз. Более того, идентифицировали даже белок, который может распознавать эти ацетилированные остатки. Так что археи, вполне вероятно, используют посттрансляционные модификации гистонов для регуляции уровня компактизации генома и экспрессии генов, как эукариоты. Более того, некоторые гистоны архей имеют не N-концевые, а C-концевые хвосты, похожие на короткий C-концевой хвост эукариотического гистона H4. В геномах некоторых архей найдены гены, кодирующие «усеченные» версии гистонов, которые, тем не менее, способны связываться с ДНК и выполнять некоторые регуляторные функции. Показано также, что посттрансляционным модификациям могут подвергаться и NAPs семейства Alba.
Несмотря на значительный прогресс в нашем понимании устройства хроматина архей, многое еще предстоит выяснить. Неизвестно, например, насколько динамичны гипернуклеосомы, и как клетки регулируют размер теоретически бесконечных гипернуклеосом? Как на структуре гипернуклеосомы отражается синтез особых форм гистонов в определенных условиях? Остается надеяться, что дальнейшее изучение «темной материи» хроматина архей поможет ответить на эти и многие другие вопросы.
Источник: БИОМОЛЕКУЛА
