
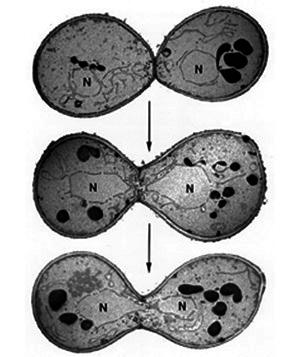
Спаривание дрожжей: две гаплоидные клетки, принадлежащие к разным полам (a и α), сливаются в одну диплоидную.
Идея о том, что половое размножение ускоряет адаптацию, неоднократно подтверждалась экспериментально. Кроме того, она имеет весомые теоретические обоснования. До сих пор, однако, ход эволюционных изменений генома в бесполых и половых популяциях не сравнивался напрямую. Американские биологи придумали методику и создали генно-модифицированные штаммы дрожжей, благодаря которым эту технически сложную задачу удалось решить. Оказалось, что при одинаковом темпе мутирования в половых популяциях за 1000 поколений закрепляется в 5 раз меньше мутаций, чем в бесполых популяциях такой же численности. Среди мутаций, закрепившихся в бесполых популяциях, встречаются вредные, тогда как в половых популяциях закрепляются только полезные мутации. Эти результаты подтверждают классические представления о том, что половое размножение ускоряет адаптацию, помогая отбору отделять полезные мутации от вредных. В бесполой популяции отбираться могут только целые геномы, поэтому каждая новая полезная мутация неразрывно связана со своим «генетическим фоном» и может распространяться только вместе с ним.
Половое размножение очень широко распространено в природе несмотря на то, что по сравнению с бесполым оно требует больших дополнительных затрат. Это значит, что половое размножение дает какие-то важные преимущества, но вот какие? Ответ на этот вопрос не так очевиден, как может показаться на первый взгляд.
Многочисленные факты, в том числе полученные в ходе эволюционных экспериментов, говорят о том, что половое размножение ускоряет адаптацию к меняющимся условиям. Этот эффект предположительно связан с тем, что половое размножение повышает эффективность закрепления отбором полезных мутаций и отбраковки вредных. При половом размножении геномы особей постоянно смешиваются и перетасовываются, а генетические варианты (аллели) оказываются всё время в разных компаниях. Это позволяет отбору работать с полезными и вредными мутациями по отдельности, закрепляя первые и отбраковывая вторые. Например, если полезная мутация возникла в геноме особи, у которой в другом гене есть вредная мутация, то в ходе полового размножения часть потомков унаследует только полезную мутацию. Они-то и будут поддержаны отбором. Кроме того, при половом размножении полезные мутации, возникшие в разных генах у разных особей, могут объединяться в одном геноме.
При бесполом (клональном) размножении отбор может работать только с целыми геномами. Поэтому полезная мутация может закрепиться лишь вместе со всем своим «генетическим фоном», включая все вредные мутации, которые есть в данном геноме. Иными словами, в бесполой популяции многие вредные мутации должны закрепляться, распространяясь за счет полезных мутаций, находящихся в других местах того же генома. Это называют «генетическим автостопом».
Кроме того, в бесполой популяции полезные мутации, возникшие в разных генах у разных особей, не могут объединиться. Вместо этого их носители начинают конкурировать и вытеснять друг друга, что ведет к безвозвратной потере большинства полезных мутаций, возникающих в популяции. Это называют конкуренцией клонов, или клональной интерференцией.
Таким образом, с одной стороны у нас есть экспериментальные данные, показывающие, что половое размножение ускоряет адаптацию, с другой — правдоподобные теоретические объяснения этого эффекта. Не хватает лишь прямых молекулярно-генетических подтверждений того, что ускорение адаптации в половых популяциях по сравнению с бесполыми действительно связано с теоретически предсказанными механизмами, такими как генетический автостоп и клональная интерференция. Чтобы получить такие подтверждения, нужно не только создать «правильные» подопытные популяции, не отличающиеся друг от друга ничем, кроме наличия или отсутствия полового размножения, но и проследить за изменениями их геномов в ходе эволюции.
Именно это и сделали биологи из Гарвардского университета (США), чья статья опубликована 24 февраля на сайте журнала Nature.
Авторы провели эволюционный эксперимент на специально изготовленных для этой цели генетически модифицированных штаммах пекарских дрожжей Saccharomyces cerevisiae. В основном эксперименте участвовало 18 подопытных популяций: 12 «бесполых», которые размножались только почкованием (митозом) и 6 «половых», которые тоже большую часть времени размножались почкованием, но после каждых 90 бесполых поколений их заставляли спариваться и производить потомство половым путем. Все популяции приспосабливались к жизни в одних и тех же условиях в течение 100 дней (примерно 1000 поколений).
Жизненный цикл дрожжей изображен на рис. 1. Гаплоидные клетки делятся на два пола: а и α. Они могут размножаться почкованием, а могут слиться в зиготу и превратиться в диплоидную клетку. Диплоидные дрожжи тоже могут размножаться почкованием, а могут перейти к споруляции и в результате мейоза превратиться в четыре гаплоидные споры, две из которых будут принадлежать к полу a, а две — к α.
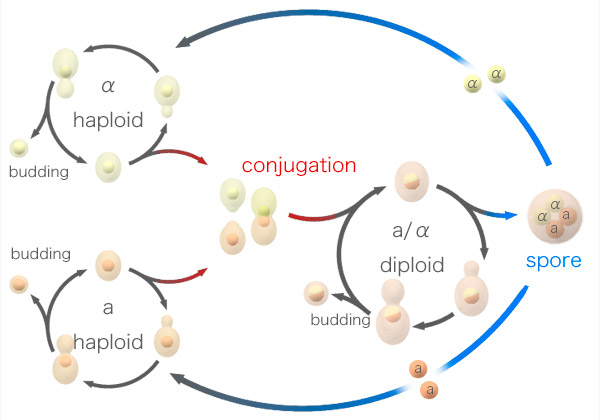
Рис. 1. Жизненный цикл дрожжей Saccharomyces cerevisiae.
В ходе эксперимента клетки бесполых популяций всегда находились в гаплоидной фазе и размножались исключительно почкованием. Чтобы они не спаривались, каждая популяция состояла из клеток только одного пола (шесть популяций a и шесть α).
Каждая половая популяция состояла из двух разнополых половинок, которые содержались раздельно в течение 90 поколений, а затем смешивались и помещались в условия, стимулирующие половой процесс. После этого отбирались диплоидные клетки, возникшие в результате спаривания, и помещались в условия, способствующие споруляции. Затем отбраковывались диплоидные клетки, не захотевшие превращаться в споры, а гаплоидные клетки, полученные из спор, разделялись по половому признаку и снова содержались раздельно в течение следующих 90 поколений.
Чтобы произвести все эти манипуляции, нужно иметь возможность быстро и эффективно отделять диплоидные клетки от гаплоидных, а также пол a от пола α. Для этого в геномы подопытных дрожжей были внесены генетические модификации. В частности, были добавлены гены устойчивости к двум антибиотикам, причем сделали это так, чтобы клетки, а оказались устойчивы только к первому антибиотику, клетки α — только ко второму, а диплоидные клетки — к обоим. Благодаря другим модификациям только гаплоидные клетки пола a умели синтезировать гистидин, только гаплоидные клетки пола α могли производить лейцин, а синтез урацила могли осуществлять гаплоидные клетки обоих полов, но не диплоидные. В результате авторы получили возможность быстро отбирать нужные клетки и контролировать чистоту опыта, попеременно используя питательные среды, содержащие (или не содержащие) урацил, гистидин, лейцин и два антибиотика в разных комбинациях.
Условия содержания бесполых и половых популяций сделали настолько одинаковыми, насколько это было возможно. Все популяции были одинаковой численности (порядка 100 000 особей), содержались в одинаковых емкостях, при одинаковой температуре и на одинаковой питательной среде (YEPD). Что касается тех манипуляций, которые проводились с половыми популяциями, чтобы побудить их к половому процессу и споруляции, то некоторые из них проводились также и с бесполыми, а влияние остальных было проверено в серии дополнительных экспериментов, которые показали, что эти манипуляции сами по себе не сказываются на ходе эволюции. Были также проведены дополнительные эволюционные эксперименты, в которых бесполые популяции тоже состояли из двух половинок, содержавшихся раздельно в течение 90 поколений, а затем смешивавшихся (но только обе половинки были одного пола). Эти тоже не повлияло на результаты.
По прошествии 1000 поколений была измерена приспособленность подопытных популяций к тем условиям, в которых они эволюционировали. Для этого исходный (предковый) штамм, помеченный флюоресцентной меткой, смешивали в равной пропорции с тестируемым штаммом и выращивали в течение 30 поколений в тех же условиях, в которых проходил основной эксперимент, а потом подсчитывали долю меченых клеток. Чем она меньше, тем выше приспособленность тестируемого штамма. Оказалось, что половые популяции адаптировались лучше бесполых: у первых приспособленность выросла на 10–15%, у вторых — лишь на 5–10%. Таким образом, в очередной раз подтвердился вывод о том, что половое размножение ускоряет адаптацию.
Чтобы разобраться в молекулярных механизмах адаптации, авторы использовали полногеномное секвенирование. Через каждые 90 поколений секвенировались выборки из четырех половых и четырех бесполых популяций. Поскольку ДНК в каждой пробе происходила не из одной, а из множества клеток и использовался мощный секвенатор Illumina HiSeq 2500, эта процедура позволила выявить многие мутации, возникавшие в популяциях в ходе эволюционного эксперимента, а также проследить изменения частоты их встречаемости со временем. Идентификация редких мутаций чревата ошибками, поэтому авторы для надежности сосредоточились только на мутациях, частота которых в данной популяции достигала 10% хотя бы в два момента времени. Таким образом, мутации, не получившие заметного распространения, были проигнорированы.
Наборы мутаций, которые в итоге были зарегистрированы как «появившиеся» в ходе эксперимента, были сопоставлены с мутациями, которые к концу эксперимента зафиксировались, то есть имелись уже у всех клеток в данной популяции (рис. 2).
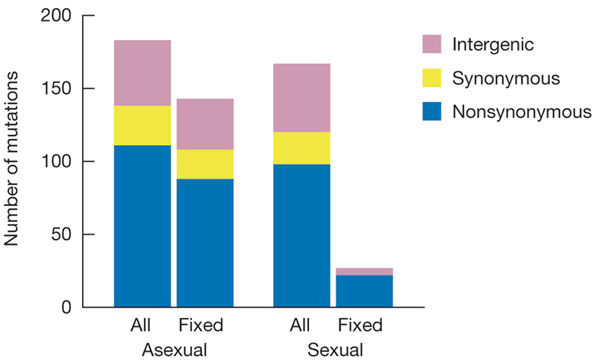
Рис. 2. Число всех появившихся мутаций (All) и тех из них, которые к концу эксперимента зафиксировались, то есть встречались у всех клеток в данной популяции (Fixed). Суммированы данные по четырем бесполым (Asexual) и четырем половым (Sexual) популяциям. Сиреневым цветом обозначены мутации в межгенных участках (intergenic), желтым — синонимичные (synonymous), синим — несинонимичные (nonsynonymous).
Оказалось, что во всех популяциях, как бесполых, так и половых, появилось примерно поровну новых мутаций (в среднем 44 на популяцию). Кроме того, среди зарегистрированных мутаций везде было примерно одинаковое соотношение несинонимичных (значимых, меняющих аминокислоту в белке), синонимичных (не меняющих белок) и межгенных (расположенных в некодирующих областях генома). Это значит, что процесс мутагенеза не различался в двух вариантах опыта.
Однако количество и состав зафиксировавшихся мутаций оказался совершенно разным в бесполых и половых популяциях. В первом случае зафиксировалось большинство (78%) зарегистрированных мутаций, в том числе 79% несинонимичных, 74% синонимичных и 78% межгенных. Иными словами, зафиксировались примерно равные доли мутаций всех трех типов. В половых популяциях всё было по-другому. Во-первых, зафиксировалось гораздо меньше мутаций (16% от общего числа зарегистрированных), во-вторых, зафиксировались в основном значимые мутации (22% от числа возникших), синонимичные не зафиксировались вовсе (0%), а межгенных закрепилось лишь 11% (рис. 2).
Этот результат отлично согласуется с идеей о том, что в половых популяциях фиксируются только полезные мутации (среди которых больше всего значимых, но есть и межгенные, находящиеся в регуляторных участках генома). Синонимичные мутации обычно нейтральны, и поэтому в половых популяциях они не фиксировались. В бесполых же популяциях вместе с немногочисленными полезными мутациями фиксировался весь их «генетический фон», включая вредные и нейтральные мутации, оказавшиеся в одном геноме с полезной. Разумеется, для этого необходимо, чтобы эффект полезной мутации перевешивал суммарные эффекты всех вредных мутаций, распространяющихся вместе с ней за счет «генетического автостопа».
Здесь необходимо пояснить, что в долгосрочной перспективе в природных популяциях фиксируется больше всего нейтральных мутаций, в том числе синонимичных. Но это происходит не за счет отбора, а за счет генетического дрейфа. Однако в рамках данного эксперимента дрейфом можно пренебречь. Для того, чтобы в популяции численностью 100 000 особей за счет дрейфа зафиксировалась новая нейтральная мутация, требуются временные интервалы порядка сотен тысяч поколений. Но эксперимент продолжался всего 1000 поколений, поэтому в подопытных популяциях за счет дрейфа ничего зафиксироваться не могло. Те мутации, которые в них зафиксировались, сделали это наверняка под действием отбора, а не дрейфа.
Таким образом, полученный результат показывает, что половое размножение помогает отбору отделять полезные мутации от всех прочих и закреплять только их, тогда как в бесполых популяциях вместе с немногочисленными полезными мутациями благодаря автостопу фиксируется много всякого мусора.
Этот вывод подтвердился при анализе динамики частоты встречаемости мутаций (рис. 3). В половых популяциях частоты разных мутаций менялись независимо друг от друга, а в бесполых большие группы функционально не связанных друг с другом мутаций меняли свою частоту синхронно. Первое соответствует индивидуальному действию отбора на отдельные участки генома, второе — клональной эволюции целых геномов. Кроме того, на рисунке 3 можно видеть, как в бесполых популяциях некоторые группы мутаций сначала быстро наращивают свою частоту, а потом их частота снижается вплоть до полного вымирания. Это — результат клональной интерференции, то есть вытеснения клонов с удачными мутациями другими клонами, обладающими еще более удачными мутациями. В половой популяции полезные мутации обоих клонов могли бы закрепиться, но в бесполой популяции они могут только конкурировать и вытеснять друг друга.

Рис. 3. Динамика частоты встречаемости новых мутаций в четырех бесполых (слева) и четырех половых популяциях. По горизонтальной оси — время в поколениях. Сплошные линии — несинонимичные мутации, крупный пунктир — синонимичные, мелкий пунктир — межгенные. Видно, что в половых популяциях мутации меняли свою частоту и фиксировались более или менее независимо друг от друга, тогда как в бесполых популяциях они делали это группами.
Чтобы еще детальнее разобраться во влиянии секса на молекулярную эволюцию, авторы попытались напрямую оценить влияние отдельных мутаций на приспособленность дрожжей. Для этого штамм, обладающий той или иной мутацией, скрещивали с предковой линией, а из гибридного потомства выделяли многочисленные клоны, содержащие эту мутацию в разных генетических контекстах. После этого следили, как растет или снижается частота мутации в ряду бесполых поколений во всех этих клонах, а результаты усредняли. Другой способ состоял в искусственном внедрении мутаций в геном предкового штамма.
Как и следовало ожидать, в каждой группе мутаций, зафиксировавшейся в бесполых популяциях (мы ведь помним, что при бесполом размножении мутации фиксируются группами), обнаружилась как минимум одна полезная мутация. Однако многие другие мутации, зафиксировавшиеся в бесполых популяциях, оказались нейтральными или вредными. Некоторые из них снижают приспособленность на 1–3%. Ясно, что они могли зафиксироваться только за счет автостопа. Идея о том, что в адаптирующихся бесполых популяциях могут закрепляться мутации со значительным вредным эффектом, высказывалась ранее, но экспериментально подтвердить ее удалось впервые.
Среди мутаций, зафиксировавшихся в половых популяциях, вредных или нейтральных обнаружить не удалось: все они оказались полезными. Правда, одна мутация, изменившая ген MET2 (кодирует фермент, участвующий в синтезе метионина), вредна в предковом генетическом контексте. Однако она полезна на фоне других мутаций, которые возникли ранее в данной подопытной линии. Зависимость полезности (и вообще фенотипического эффекта) мутации от генетического контекста называют эпистазом. Ранее уже было показано, что в эволюции дрожжей эпистаз играет некоторую роль, правда далеко не такую важную, как у бактерий.
Данное исследование относится к числу работ, подтверждающих и раскрывающих на более глубоком, молекулярном уровне старые, классические эволюционные идеи. В данном случае блестяще подтвердилась идея о том, что половое размножение ускоряет адаптацию, помогая отбору отделять полезные мутации от вредных. Благодаря половому размножению блокируются такие неблагоприятные для популяции процессы, как генетический автостоп, помогающий фиксироваться вредным мутациям, и клональная интерференция, обрекающая на гибель большинство вновь возникающих полезных мутаций.
Источник: ЭЛЕМЕНТЫ БОЛЬШОЙ НАУКИ
