
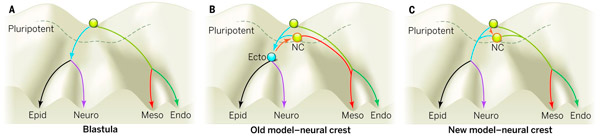
Рис. 1. А. «Эпигенетический ландшафт» по Конраду Уоддингтону. Эмбриональные клетки изображаются в виде шариков, катящихся по системе ветвящихся желобов от начальной точки, соответствующей плюрипотентному к своей конечной «цели» — необратимому превращению в ту или иную специализированную клетку. По мере движения клетка последовательно теряет альтернативные возможности развития, становясь клеткой эпидермиса, нейроэктодермы, мезодермы или энтодермы. B. Ранее считалось, что плюрипотентные клетки нервного гребня (NC, neural crest) происходят от дифференцированных эктодермальных клеток и приобретают свою плюрипотентность вторично. Это противоречило классической схеме Уоддингтона, предполагая обратное движение клетки вверх по склону «ландшафта развития». C. Новые данные показывают, что клетки нервного гребня просто катятся медленнее остальных: они сохраняют исходную плюрипотентность, характерную для клеток раннего эмбриона, в то время как остальные клетки ее утрачивают.
Нервный гребень — загадочная эмбриональная структура, имеющаяся только у позвоночных. Его часто называют «четвёртым зародышевым листком», помещая в один ряд с классическими экто-, эндо- и мезодермой, хотя формально он является производным эктодермы. До сих пор оставалось не вполне ясным, как удается клеткам нервного гребня, расползаясь по эмбриону, давать начало различным тканям и типам клеток, которые у всех животных, кроме позвоночных, формируются из мезодермы. Американские ученые приблизились к решению этой эмбриологической загадки, показав, что плюрипотентность (способность превращаться в разные типы клеток) поддерживается в клетках нервного гребня благодаря работе тех же регуляторных генов, что и в эмбриональных стволовых клетках. По-видимому, клетки нервного гребня не приобретают плюрипотентность вторично, как предполагалось ранее, а наследуют ее напрямую от плюрипотентных клеток раннего эмбриона.
Развитие животных начинается с одной тотипотентной (способной дать начало всем тканям и частям организма) клетки — зиготы, или оплодотворенной яйцеклетки. Затем, по мере развития зародыша, проиcходит последовательная дифференцировка клеток, сопровождающаяся утратой альтернативных возможностей развития.
На ранних этапах (на стадии бластулы или бластоцисты) в эмбрионе присутствуют так называемые плюрипотентные клетки, способные дать начало всем тканям и типам клеток, за исключением некоторых специфических зародышевых структур, которые не войдут в состав взрослого организма. У млекопитающих плюрипотентными являются клетки внутренней клеточной массы на стадии бластоцисты; именно из них получают лабораторные культуры эмбриональных стволовых клеток (ЭСК). У шпорцевой лягушки Xenopus, на которой выполнено обсуждаемое исследование, плюрипотентными являются клетки анимального полюса бластулы.
У всех билатерально-симметричных животных плюрипотентные клетки в ходе развития подразделяются (дифференцируются) на три зародышевых листка: наружную эктодерму (из нее затем разовьются эпидермис и нервная система), внутреннюю энтодерму (пищеварительный тракт и его производные) и находящуюся посередине мезодерму, из которой разовьется всё остальное: мышцы, кости (у кого они есть), выделительная система и т. д. Как правило, подразделение на три зародышевых листка необратимо. Если клетка стала эктодермальной, из нее уже нельзя вырастить мышцы или кости, а превращение клетки в мезодермальную лишает ее способности давать начало клеткам эпидермиса и нейронам мозга.
Нервный гребень позвоночных — исключение из этого общего правила. Он образуется, когда три зародышевых листка давно уже сформированы, причем образуется он как часть эктодермы (по краям нервной пластинки; подробнее см. в вышеупомянутой новости). Несмотря на свою очевидную эктодермальную природу, клетки нервного гребня дают начало многим тканям и типам клеток, в том числе таким, которые у всех животных, кроме позвоночных, формируются из мезодермы. В частности, из клеток нервного гребня образуется значительная часть костных и хрящевых структур головы, включая челюсти и зубы.
Итак, клетки нервного гребня обладают плюрипотентностью, сопоставимой с плюрипотентностью эмбриональных стволовых клеток. Причем происходит всё это в то время, когда остальные клетки эмбриона, включая прилегающие к нервному гребню участки эктодермы, давно уже распрощались с плюрипотентностью, выбрали свою судьбу и стали кто клетками эпидермиса, кто клетками нейроэктодермы (нервной пластинки — будущей центральной нервной системы), кто чем-то еще.
Природа плюрипотентности клеток нервного гребня — одна из интригующих эмбриологических загадок, ждущих своего решения. Она противоречит общему правилу, согласно которому в ходе развития эмбриона происходит прогрессирующее и необратимое сужение онтогенетического потенциала клеток (рис. 1). Многие ученые предполагали, что клетки нервного гребня, будучи производными уже утративших плюрипотентность клеток эктодермы, приобретают свою плюрипотентность вторично, примерно так, как другие популяции эмбриональных клеток приобретают те или иные новые свойства — в результате действия каких-то внешних сигналов и сигнально-регуляторных генных каскадов, реагирующих на эти сигналы и меняющих экспрессию тех или иных генов. Однако поиски таких механизмов повторного приобретения плюрипотентности оказались тщетными.
Другое возможное объяснение состоит в том, что клетки нервного гребня не приобретают плюрипотентность заново, а каким-то образом ухитряются сохранить широкие возможности, доставшиеся им по наследству от ранних плюрипотентных клеток бластулы (в то время как все остальные клетки зародыша эти возможности утрачивают). Некоторые факты свидетельствуют в пользу этой второй версии: было показано, что регуляторные гены Myc, Id3 и Sox5 работают как в формирующихся клетках нервного гребня, так и в плюрипотентных клетках бластулы. Вероятно, они играют какую-то роль в поддержании плюрипотентности тех и других.
Американские эмбриологи провели целенаправленную проверку гипотезы о сохранении клетками нервного гребня первичной плюрипотентности. Для этого они сравнили экспрессию регуляторных генов в первичных плюрипотентных клетках на стадии бластулы и в клетках формирующегося нервного гребня на более поздних стадиях (гаструлы и нейрулы).
В результате удалось существенно расширить список регуляторных генов, о которых ранее было известно, что они нужны для развития нервного гребня, и которые, как выяснилось, работают также и в плюрипотентных клетках бластулы шпорцевой лягушки. Кроме упомянутых выше, к ним относятся гены TF-AP2, Ets1, FoxD3 и Snail1 (последние два гена работают также и в эмбриональных стволовых клетках мыши). И наоборот, многие гены, о которых ранее было известно, что они важны для поддержания плюрипотентности клеток бластулы, как выяснилось, активны также и в клетках формирующегося нервного гребня (Oct60, Sox3, Vent2 и другие).
Как показали дальнейшие эксперименты, эти гены, скорее всего, являются универсальными «факторами плюрипотентности». В частности, подтвердилось, что гены из первой группы («гены нервного гребня», работающие также и в плюрипотентных клетках бластулы) действительно необходимы для поддержания плюрипотентности последних. Их отключение ведет к прекращению экспрессии и других «факторов плюрипотентности». В результате плюрипотентность утрачивается. Например, если отключить в клетках бластулы ген Snail1, клетки теряют способность превращаться в клетки мезодермы и энтодермы. Они перестают реагировать на сигнальные белки (Nodal и activin), которые в норме индуцируют их превращение в мезодермальные или энтодермальные клетки.
Дальнейшие эксперименты с эмбрионами и их фрагментами (эксплантами) показали, что если искусственно превратить плюрипотентные клетки анимального полюса бластулы в клетки нервного гребня (для этого в клетках нужно одновременно активировать ген Snail2 и сигнальный путь Wnt), то такие клетки сохраняют плюрипотентность намного дольше обычного. Это проявляется в том, что они в ответ на соответствующие химические сигналы успешно превращаются не только в мезодермальные клетки (это нормально для клеток нервного гребня), но и в энтодермальные (чего при нормальном развитии зародыша клетки нервного гребня вроде бы не делают). Более того, выяснилось, что такими же свойствами обладают и обычные (а не полученные искусственно) клетки нервного гребня. В частности, если на них воздействовать высокой концентрацией сигнального белка активина, они начинают экспрессировать энтодермальные маркеры Endodermin и Sox17, то есть фактически превращаются в энтодерму. Другими словами, когда ученые культивировали в лаборатории клетки нервного гребня, оказалось возможным запустить их не только по мезодермальному пути развития (на котором можно получить кости и хрящи), но, что неожиданно, даже по энтодермальному пути.
Полученные результаты показывают, что клетки нервного гребня, по всей видимости, не восстанавливают плюрипотентность, побыв какое-то время клетками эктодермы с ограниченным потенциалом, а сохраняют исходный широкий набор возможных путей развития, наследуя его напрямую от своих предшественников — клеток анимального полюса бластулы (рис. 2). Плюрипотентность в обоих случаях поддерживается благодаря активности одних и тех же регуляторных генов. Авторы образно называют обнаруженное явление «клеточной неотенией» (неотения — сохранение на поздних стадиях развития признаков, характерных для ранних стадий). Клетки нервного гребня сохраняют «детскую» широту возможностей в то время, когда все остальные клетки зародыша уже давно ее утратили, став «узкими специалистами».
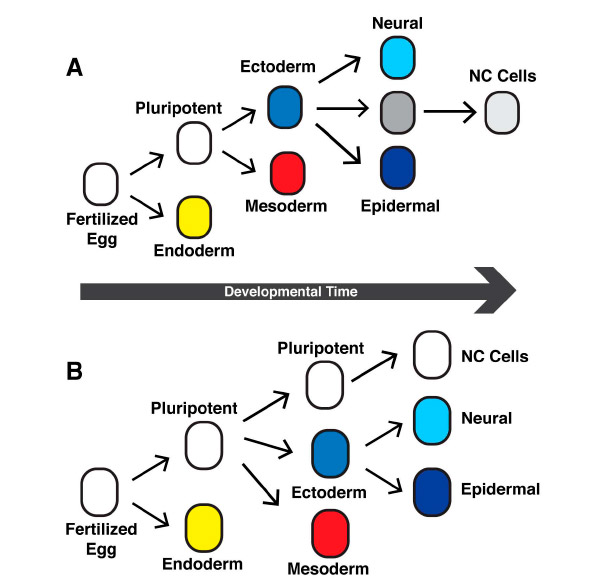
Рис. 2. Старая (вверху) и новая (внизу) схемы дифференцировки клеток в ходе развития зародыша шпорцевой лягушки. По новым данным, клетки нервного гребня (NC cells) не приобретают плюрипотентность вторично, а наследуют ее напрямую от плюрипотентных клеток раннего эмбриона.
Данное исследование — важный шаг к пониманию происхождения и эволюционного успеха позвоночных. Ведь из клеток нервного гребня формируются многие морфологические структуры, имеющие к этому успеху самое прямое отношение — такие, например, как череп, челюсти и зубы.
Источник: ЭЛЕМЕНТЫ БОЛЬШОЙ НАУКИ
