
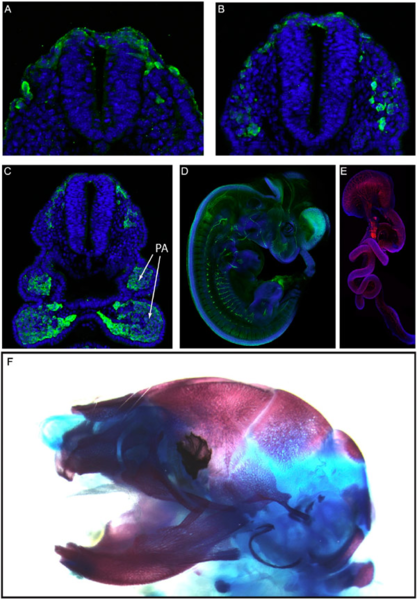
Рис. 1. Миграция и дифференциация нервного гребня у мыши. A–C — поперечные разрезы через зародыш вскоре после образования нервной трубки. Клетки нервного гребня окрашены зеленым, другие клетки — синим. A — начало миграции нервного гребня, B — продолжение миграции: потоки клеток нервного гребня пробиваются сквозь мезодерму; C — многие клетки нервного гребня уже достигли своего окончательного положения, а именно — зачатков жаберных дуг (PA, pharyngeal arches). D — фотография более позднего зародыша целиком; в зеленый цвет окрашены производные нервного гребня, относящиеся к периферической нервной системе. E — кишечник с расположенными в нем сплетениями метасимпатической нервной системы, которая тоже является производным нервного гребня; здесь она окрашена красным. F — череп, в основном образованный скелетными производными нервного гребня: кость (красная) и хрящ (синий).
У позвоночных животных есть особый эмбриональный зачаток, называемый нервным гребнем (он расположен рядом с нервной трубкой). Из клеток нервного гребня образуется удивительно много разных структур, от некоторых нервных узлов до большей части черепа. Многие современные ученые считают нервный гребень четвертым зародышевым листком, наряду с эктодермой, энтодермой и мезодермой. У ближайших родственников позвоночных — оболочников — есть группа зародышевых клеток, близкая по свойствам к нервному гребню, которая дифференцируется в пигментные клетки покровов. Вероятно, эта группа клеток сохранилась и у позвоночных, значительно расширив набор путей своей дифференцировки. Кроме того, у позвоночных появились новые регуляторные гены с экспрессией, специфичной для нервного гребня; это было облегчено тем, что в их эволюции произошла полногеномная дупликация. Таким образом, две уникальные особенности подтипа позвоночных — полногеномная дупликация и присутствие «четвертого зародышевого листка» — наверняка связаны между собой.
Можно ли свести устройство всех животных к единой схеме? Простого ответа на этот вопрос нет. Все зависит от детальности требуемой схемы и от того, как именно мы ее собираемся использовать. Тем не менее вопрос о наличии у животных «единого плана строения» рассматривался в классической зоологии как важнейший, и между сторонниками разных ответов на него бывали грандиозные споры. И правда, вопрос этот важен — хотя бы потому, что любая наука стремится описывать свои объекты по общему для всех шаблону, а «единый план строения» как раз и мог бы предоставить такой шаблон.
В середине XIX века эмбриология подарила эволюционной науке ценное обобщение, позволившее, по крайней мере, сравнивать сколь угодно разных животных между собой. Было установлено, что зародыш любого (или почти любого) животного, достигнув определенной стадии, делится на устойчивые слои клеток, которые называются зародышевыми листками. Всего зародышевых листков три: эктодерма (наружный), энтодерма (внутренний) и мезодерма (средний). Из эктодермы образуется кожный покров (эпидермис) и нервная система. Из энтодермы образуется кишечник — точнее, пищеварительный тракт — и органы, развивающиеся как его выросты, например печень. Из мезодермы, как правило, образуются опорно-двигательная, кровеносная и выделительная системы.
У некоторых животных (например, у гидроидных полипов, к которым относится пресноводная гидра) есть эктодерма и энтодерма, но мезодермы нет. У двусторонне-симметричных животных, к которым относимся и мы, есть все три зародышевых листка. Животных с двумя зародышевыми листками называют двуслойными (диплобластами), животных с тремя зародышевыми листками — трехслойными (триплобластами).
Автор известного курса общей эмбриологии Б. П. Токин назвал теорию зародышевых листков «самым крупным морфологическим обобщением за всю историю эмбриологии». К рубежу XIX–XX веков эта теория стала общепринятой. Более того, сложилось своеобразное представление о «святости» зародышевых листков, границы которых считались непоколебимыми. Если некоторый орган образуется из одного зародышевого листка, он никогда, ни у какого организма не может образоваться из другого.
Но, как часто бывает, живая природа оказалась объемнее академических схем. В данном случае это выяснилось быстро. В 1893 году американская исследовательница-эмбриолог Джулия Платт обнаружила, что некоторые хрящи жаберного аппарата позвоночных развиваются не из мезодермы (как следовало бы ожидать по классической теории зародышевых листков), а из эктодермы. Джулия Платт сделала целую серию работ по прослеживанию судьбы эктодермальных клеток, образующих хрящи. Ее результаты были подтверждены несколькими другими эмбриологами. Но широкого признания это открытие не нашло, в основном из-за чисто догматических сомнений: хрящам «положено» развиваться из мезодермы — значит, развиваться из эктодермы они не могут, и всё тут! Джулия Платт даже не получила постоянной ставки в университете, после чего решила вообще оставить науку. Она занялась общественной деятельностью, стала заметным в штате Калифорния политиком, много сделала для охраны природы, так что человечество в целом тут, может, и не пострадало. Но вот особое происхождение жаберных хрящей стало общепринятым фактом только в конце 1940-х годов, после очень тонких опытов шведского эмбриолога Свена Хёрстадиуса, усомниться в результатах которых было уже трудно.
Казалось бы, какое значение для нашего мировоззрения может иметь вопрос о том, из каких именно зародышевых клеток формируются жаберные дуги тритона или акулы? Не мелочь ли это? Нет, не мелочь. Потянув, как за ниточку, за данные Платт и Хёрстадиуса, мы оказываемся перед серьезной макроэволюционной проблемой.
Мы уже знаем, что эктодерма — самый внешний из трех зародышевых листков. У позвоночных она делится на две части: (1) покровная эктодерма и (2) нейроэктодерма. Из покровной эктодермы образуется эпидермис, из нейроэктодермы — центральная нервная система. Покровная эктодерма, естественно, одевает тело будущего животного снаружи. Что касается нейроэктодермы, то она сначала представляет собой расположенную на будущей спине нервную пластинку, которая потом погружается, сворачивается и замыкается в нервную трубку. Эта трубка и становится центральной нервной системой, то есть мозгом (спинным и головным).
На самой границе нейроэктодермы и покровной эктодермы у позвоночных находится группа клеток, называемая нервным валиком, или нервным гребнем. Клетки нервного гребня не входят ни в состав нервной трубки, ни в состав эпидермиса. Зато они способны расползаться по всему организму, мигрируя, как амебы, с помощью ложноножек. Именно судьбу клеток нервного гребня и изучала Джулия Платт. Действительно, из них формируются многочисленные структуры, далеко не только нервные. Свен Хёрстадиус в свое время показал, что если у зародыша хвостатой амфибии микрохирургически удалить нервный гребень в передней трети тела, то у него нормально развивается затылок, нормально развиваются ушные капсулы — а всего остального черепа просто нет. Ни большая часть мозговой коробки, ни капсулы органов обоняния, ни челюсти не развиваются без вклада клеток нервного гребня (рис. 2).
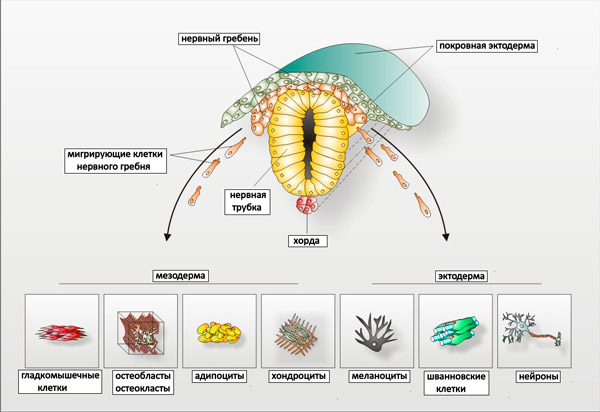
Рис. 2. Схема зародыша позвоночного на стадии миграции клеток нервного гребня (поперечный разрез). Внизу в условных областях, принадлежащих эктодерме и мезодерме, показаны типы клеток, которые могут дифференцироваться не из этих тканей, а из нервного гребня. Остеобласты и остеокласты — клетки кости, хондроциты — клетки хряща; остальные пояснения см. в тексте.
Вот список (наверняка неполный) производных нервного гребня у позвоночных:
- Нервные узлы спинных корешков спинномозговых нервов (часто их называют просто спинальными ганглиями).
- Нервные узлы вегетативной нервной системы (симпатической, парасимпатической и метасимпатической).
- Мозговое вещество надпочечников.
- Шванновские клетки, образующие оболочку отростков нейронов.
- Внутренняя выстилка (эндотелий) и гладкомышечный слой некоторых сосудов, в том числе аорты.
- Ресничные мышцы, сужающие и расширяющие зрачок.
- Одонтобласты — клетки, выделяющие дентин, твердое вещество зубов.
- Пигментные клетки покровов: эритрофоры (красные), ксантофоры (желтые), иридофоры (отражающие), меланофоры и меланоциты (черные).
- Часть адипоцитов — клеток жировой ткани.
- Парафолликулярные клетки щитовидной железы, выделяющие гормон кальцитонин.
- Хрящи и кости черепа, в первую очередь его висцерального (глоточного) отдела, в который входят не только жаберные дуги, но и челюсти.
Богатый перечень, не правда ли? Ну, спинномозговые ганглии — это неудивительно: они расположены как раз примерно на месте нервного гребня, клеткам которого в данном случае даже не приходится совершать миграцию. Вегетативные ганглии — тоже ничего удивительного. Они расположены гораздо дальше от спинного мозга, но, в конце концов, это часть нервной системы. И мозговое вещество надпочечников — это фактически вегетативный ганглий, только преобразованный. И шванновские клетки — часть нервной ткани. Но дальше-то в списке идут структуры, не имеющие к нервной системе никакого отношения, притом разнообразные и многочисленные. У человека есть и болезни, вызываемые аномалиями производных нервного гребня, — нейрокристопатии.
Исключительно важен последний пункт списка: череп! Из нервного гребня образуется, собственно, большая его часть (кроме слухового отдела и затылка). Между тем весь остальной скелет — позвоночник, скелет конечностей — образуется из мезодермы. Классическая концепция, согласно которой однотипные органы не должны развиваться из разных зародышевых листков, тут явно дала сбой.
Еще один важный момент: весь список производных нервного гребня относится не к хордовым, а именно к позвоночным. Кроме позвоночных в тип хордовых входят еще две современные группы животных: оболочники и ланцетники. Так вот у них нервный гребень не выражен. Это уникальный признак подтипа позвоночных.
Что же такое нервный гребень? Если это часть эктодермы, то какая-то уж слишком необычная. В 2000 году канадский эмбриолог Брайан Холл предложил считать нервный гребень не чем иным, как отдельным — четвертым — зародышевым листком. Эта трактовка быстро распространилась в научной литературе, где нервный гребень сейчас вообще является популярной темой. Получается, что позвоночные — единственные четырехслойные животные (квадробласты).
Четвертый зародышевый листок — такая же важная особенность позвоночных, как, например, случившаяся в начале их эволюции полногеномная дупликация. Но как он возник? Американские биологи Уильям Муньос и Пол Трэйнор опубликовали статью о современном состоянии этой проблемы (рис. 1). Пол Трэйнор — видный эмбриолог позвоночных, уже много лет специализирующийся как раз на нервном гребне, так что обзор, подписанный им, точно заслуживает внимания.
По современным данным, от эволюционного древа хордовых первой отошла веточка, ведущая к ланцетнику. Оболочники и позвоночные — более близкие родственники; вместе они образуют группу, которая называется Olfactores («животные с органом обоняния»). Раз ланцетник представляет более древнюю ветвь, то у него можно ожидать более древних признаков. Действительно, никаких близких аналогов клеток нервного гребня у ланцетника не найдено. Большинства органов и тканей, которые у позвоночных образуются из материала нервного гребня, в его теле просто нет. Существует одно серьезное исключение: волокна чувствительных спинномозговых нервов ланцетника окружены вспомогательными (глиальными) клетками, очень похожими на шванновские клетки позвоночных. Шванновские клетки — важнейшие производные нервного гребня. Но их аналоги у ланцетника образуются из обычной нейроэктодермы, то есть из материала нервной трубки. Этот пример только подтверждает: никакого нервного гребня у ланцетника нет.
С оболочниками дело обстоит сложнее и интереснее. У асцидии Ciona intestinalis (вполне типичный и хорошо изученный оболочник) аналоги производных нервного гребня есть — это пигментные клетки, содержащие меланин. И их эмбриональный источник расположен как раз «где надо»: на границе нервной пластинки и покровной эктодермы. Особенности индивидуального развития асцидии позволяют проследить судьбу этих клеток очень точно. Прежде чем занять свое место в покровах, они совершают долгую миграцию (иногда сквозь рыхлую мезодерму, а иногда между мезодермой и эпидермисом); все это очень похоже на поведение клеток типичного нервного гребня. Более того, в предшественниках пигментных клеток асцидии экспрессируется антиген HNK-1, специфичный для клеток нервного гребня позвоночных, вплоть до птиц и млекопитающих.
«Нервный гребень» асцидии происходит от определенного бластомера (то есть от определенной клетки раннего зародыша; для асцидии составлена карта раннего развития, где все бластомеры пронумерованы). Интересно, что пигментными клетками становятся не все потомки этого бластомера. Некоторые из них входят в состав мезодермы и могут, например, становиться клетками крови или мышцами стенки тела. Связь нервного гребня и мезодермы изучена пока недостаточно подробно, но она наверняка не случайна. Похоже, здесь мы прикоснулись к довольно тонкому и глубокому эволюционному механизму. У большинства животных пигментные клетки развиваются именно из мезодермы. Скорее всего, так было и у предков асцидии. Затем, в процессе эволюции хордовых, возникающий нервный гребень «перехватил» у мезодермы путь дифференцировки пигментных клеток, начав формировать их из себя. У позвоночных этот процесс продолжился: нервный гребень «перехватил» пути дифференцировки еще и таких традиционно мезодермальных тканей, как хрящ, кость, жировая ткань и гладкие мышцы, причем во всех этих случаях — только частично.
Именно так мог бы проявляться меторизис — процесс изменения границы зародышевых листков, когда один из них частично замещает другой. Это понятие ввел в 1908 году профессор Петербургского (впоследствии Петроградского) университета, академик Владимир Михайлович Шимкевич. Но Шимкевич не знал, что путем меторизиса может образоваться целый новый зародышевый листок. У позвоночных, получается, произошло именно это. Вот чем уникален их план строения.
Скелетная ткань, которая у всех известных нам животных развивается исключительно из нервного гребня — это дентин. К счастью, дентин очень тверд, и он отлично сохраняется в ископаемом состоянии. Например, мы знаем, что представители одной из самых древних групп бесчелюстных позвоночных — Pteraspidomorphi — были буквально закованы в броню из дентина (рис. 3). Видимо, это можно рассматривать как документальное свидетельство того, что нервный гребень у них уже был полностью развит. Но скорее всего, он возник еще раньше.
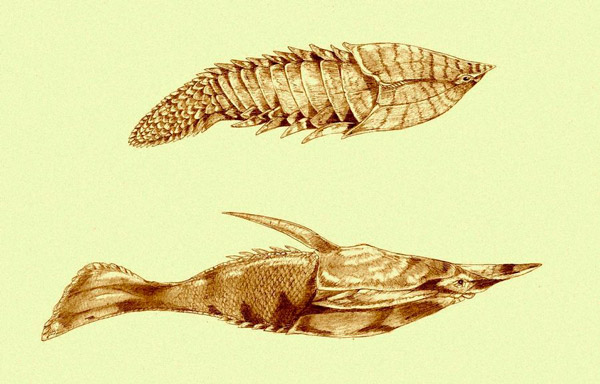
Рис. 3. Два ископаемых панцирных бесчелюстных из группы Pteraspidomorphi: Anglaspis (сверху) и Pteraspis (снизу). Великолепно развитая покровная «броня» у них состоит из дентина, который, скорее всего, и тогда был производным нервного гребня.
Остается еще один интригующий вопрос. Связаны ли между собой два уникальных признака позвоночных: четвертый зародышевый листок и полногеномная дупликация?
Да, такая связь скорее всего есть. Это можно показать на некоторых примерах, несмотря на то, что система генов, управляющих развитием нервного гребня, изучена пока не очень полно. Считается общепризнанным, что в начале эволюции позвоночных произошло подряд два события полногеномной дупликации (whole-genome duplication event, WGD). Дупликация, то есть удвоение всего генома, не может не привести к появлению дополнительных копий генов, в том числе и контролирующих индивидуальное развитие. Пример такого гена — ген FoxD, относящийся к крупному генному семейству Fox. У ланцетника этот ген один. Область его экспрессии включает некоторые участки нервной трубки, а также осевую мезодерму. У асцидии ген FoxD тоже один, поскольку никакой полногеномной дупликации у оболочников не было. Но у асцидии, в отличие от ланцетника, есть зачаток нервного гребня. Ген FoxD экспрессируется и в нем тоже. А у позвоночных генов FoxD становится несколько, и в клетках нервного гребня экспрессируется только один из них — ген FoxD3. Это — разделение функций, типичное для последствий дупликации. Есть идея, что любая дупликация сама по себе «побуждает» новые копии гена по возможности разделять между собой задачи, чтобы в генной сети не возникало сбоев из-за дублирования.
С другой стороны, можно сказать, что дупликация дала геному позвоночных дополнительные степени свободы, которые пригодились, в частности, при создании нового зародышевого листка. Ведь у асцидии такого разнообразия производных нервного гребня нет и отдаленно; у них это рядовой мелкий зачаток, обеспечивающий формирование единственного типа клеток. У позвоночных этот зачаток «взбесился», захватив огромное количество разных путей дифференцировки вместе с типами клеток, к которым эти пути ведут. А увеличение числа генов явно послужило тут предпосылкой.
В свете этих данных старое наивное представление, что позвоночные устроены сложнее всех других животных, начинает, как ни странно, выглядеть верным. Полногеномная дупликация и новый зародышевый листок — весомые объективные показатели сложности. Еще одним подобным показателем может быть, например, количество регуляторных микроРНК. Но пример с нервным гребнем даже ярче.
Источник: ЭЛЕМЕНТЫ БОЛЬШОЙ НАУКИ
